管胞
管胞(英語:Tracheid),又稱假導管[1],是維管束植物木質部中一種長形、木質化的細胞,其名稱最早於1863年由德國植物學家卡爾·古斯塔夫·薩尼歐使用,來自德文的Tracheide[2]。管胞的細胞壁上常有壁孔(又稱紋孔或導孔)或飾紋,成熟後不具原生質體,主要功能為進行水分與無機鹽的運輸,以及提供樹木的結構支撐。被子植物木質部中除管胞外,還有另一種稱為導管的輸水結構,管胞的管徑小於導管,且不具有導管細胞間穿孔板(perforation plate)的結構;大多數蕨類植物與裸子植物木質部中則不具有導管,只有管胞負責水分的運輸,松柏等裸子植物的木材(軟木)即主要是由管胞組成。
.png)
管胞在演化上可能是單一起源的構造,化石紀錄顯示早期植物的管胞有S型、G型與P型三種,其中前兩者的管壁木質化的區域呈帶狀,並有孔洞以利水分在細胞間的運輸;後者則具有類似現生植物管胞的壁孔,後來亦有結構更複雜的壁孔出現,例如許多裸子植物管胞上的具緣壁孔(bordered pit,又稱有簷壁孔、具緣紋孔或重緣紋孔),使植物在管胞間運送水分的同時,得以降低氣穴栓塞(embolization)在管胞間傳遞的風險。
形態 编辑

管胞是維管束植物木質部中的一種長形細胞,其長度約為直徑的100倍,平均直徑為20-60微米[註 1],平均長度為2-6毫米[4]。管胞的功能為負責輸送水分,大部分被子植物、買麻藤與少數蕨類植物[註 2]木質部中除了管胞外,還有另一種亦由長形細胞組成、負責輸送水分與無機鹽的結構,稱為導管,大多數裸子植物[註 3]與蕨類植物則不具有導管,僅仰賴管胞進行水分運輸[8],松柏等裸子植物的木材(軟木)即有高達九成的體積是由管胞組成[4]。導管的直徑可達500微米,輸水效率大大高於管胞[註 4][10],管胞雖輸水效率不如導管,但因管徑比導管窄而對擠壓的抗性較高,對植物的結構支撐十分重要,另外相較於導管細胞間以穿孔板(perforation plate)上下相連,形成連貫的導管,管胞細胞的兩端不具有穿孔版,且形狀較導管細胞為尖[11],水分需經由細胞間相互對準的壁孔在不同管胞間垂直流動[12]。
管胞與導管細胞成熟後均會形成木質化的次生細胞壁,且內部的原生質體經細胞凋亡而降解消失,即成為死細胞[13],少數區域不進行木質化而僅具有初生細胞壁與中膠層,即為水分可通透的壁孔[8]。導管細胞一般自維管束形成層分化後數天即完成發育並凋亡,管胞與纖維細胞的發育過程則較長,約一個月才完成凋亡過程,顯示管胞的凋亡機理可能與纖維細胞較為接近,導管細胞則另外演化出新的凋亡機理[14]。
管胞依木質化加厚的紋路可分為環紋(annular)、螺紋(helical)、梯紋(scalariform)、網紋(reticulate)與孔紋(pitted)等類型[15],初生木質部(primary xylem)中,原生木質部(protoxylem)的管胞常為環紋或螺紋,因木質化部分較少而尚有延展性,後生木質部(metaxylem)的管胞多為梯紋、網紋或孔紋;次生木質部(secondary xylem)中的管胞則大多為孔紋,即具有壁孔,且除壁孔之外的區域均已木質化,有些植物的次生木質部中也具有梯紋導管[16]。
壁孔 编辑
壁孔是管胞與導管壁上未經木質化的部分,只由初生細胞壁與中膠層組成,故可使水分通透,經常在相鄰細胞間成對出現,以便於水分在輸水細胞間的流動[8]。除了最簡單的壁孔外,部分植物的壁孔還有一些特化的結構,如常見於裸子植物管胞的具緣壁孔(bordered pit)與部分買麻藤和部分被子植物管胞或導管中的附物壁孔(vestured pit)[註 5]。
具緣壁孔 编辑
裸子植物的管胞周圍常有具緣壁孔(bordered pit),此種壁孔為細胞壁木質化的部分朝細胞內的方向延伸所形成的穹頂狀構造,其中央因膜木質化形成壁孔閥(torus,又稱紋孔閥)而不可通透,周圍的塞缘(margo)則可讓水分自由通過;被子植物的導管則較少有具緣壁孔。當管胞發生氣穴栓塞(embolization)時,壁孔塞受到擠壓,可延展到一側的壁孔緣(pit border)上將壁孔堵住,以避免氣穴進入相鄰的管胞中[8]。


特化 编辑
_(20309933680).jpg)
- 髓線中橫向生長的髓線管胞
- 髓線中的薄壁細胞
- 髓線
- 一般管胞上的壁孔
部分植物具有特殊型態的管胞[19]。裸子植物中,落葉松、花旗松等針葉木具有隔膜管胞(strand tracheid),為被縱向分割成數個細胞的管胞,其長度短於一般管胞,且壁的厚度不若一般管胞厚,壁孔大小也較小,形態介於管胞與縱向薄壁細胞(axial parenchyma)之間[20][21][22];松屬、雲杉屬、落葉松屬與黃杉屬植物的髓線中具有橫向生長的髓線管胞(ray tracheid),不同於一般管胞的縱向生長,且壁孔常較一般管胞小,壁上常有鋸齒狀的增厚紋路[20][23],髓線管胞通常位於髓線的邊緣,但也有髓線完全由髓線管胞組成的情形[24]:20;部分科達樹屬、貝殼杉屬、南洋杉屬、松屬、冷杉屬具有樹脂管胞(resinous tracheid),為內部具有樹脂的管胞,常出現在髓線周圍,樹脂可能為髓線中的薄壁細胞所合成,並貯存於此類管胞中,樹脂可阻斷管胞中水分的縱向運輸,美國植物學家大衛·P·裴哈洛認為此類管胞中的樹脂與被子植物的侵填體相似,可能具有類似功能[25];部分被子植物具有維管管胞(vascular tracheid)、環管管胞(vasicentric tracheid)或纖維管胞(fiber tracheid),維管管胞與纖維管胞可能分別為演化上介於管胞與導管細胞和木材纖維間的構造,環管管胞則可能是退化的導管,因寬度很窄而不具有穿孔板[24]:22,維管管胞與環管管胞可在鄰近的導管被氣穴堵塞時進行水分運輸,纖維管胞則主司支撐功能[26]。
演化 编辑
在植物演化出管胞之前,阿格劳蕨與羊角蕨等前維管束植物(protracheophytes)可能使用管徑較窄且壁面平滑的管狀細胞來運輸水分,其表面有原生質絲衍生的微小孔洞,雖未有木質化,但不排除也有與木質素相似的多酚類化合物以加強其結構。管胞的結構因具有木質化的次生細胞壁而較原始的輸水構造堅固許多,但因木質素為疏水材料,會提高輸水的阻力[18]。管胞(維管束植物)在演化上可能是單次起源的,從化石紀錄中可見早期的維管束植物有S型、G型與P型三種形態的管胞,其中前兩者的管壁紋路為帶狀(banded),出現於泥盆紀早期;後者則為孔狀(pitted),出現於泥盆紀中期[18]。萊尼蕨的管胞為S型(S-type,得名自Sennicaulis屬),壁上有環紋或螺紋的帶狀增厚結構,大致可分為兩層,外層為海綿狀的結構,孔洞部分可能是未經木質化而沒有被保存成化石的組織,內部則有一薄層的木質化結構,其中有均勻散播的許多微小孔洞,可能可使水分通透[18];石松類(包括工蕨類)的管胞為G型(G-type,得名自Gosslingia屬),亦有環紋或螺紋的帶狀增厚結構,且加厚部分常有中空,可能為未被保存成化石的組織,管壁有較大(4微米)的孔洞,且只分布於管壁未加厚處,而非均勻分布;真葉植物的管胞則為P型(P-type,得名自裸蕨屬Psilophyton),壁的增厚部分亦常為中空(即可能是未經木質化而沒有被保存成化石的組織),增厚部分之間有梯狀分佈的壁孔,已具有包括壁孔腔(pit chamber)與壁孔膜(pit chamber)的完整結構[27]。S型與G型的管胞與現生植物原生木質部(protoxylem)的管胞相似,P型則與後生木質部(metaxylem)與次生木質部(secondary xylem)的管胞較接近[18][28]。
輸水組織輸送水分的效率與管徑的四次方成正比[9],管胞經近4000萬年的演化,管徑漸增,至泥盆紀晚期時已達到接近現生植物管胞的管徑(約80微米),其直徑沒有再繼續增加的原因,可能是受到水分經壁孔在細胞間輸送的效率所限制,壁孔的數目與管胞管壁的表面積有關,而表面積只與管徑與管長的一次方成正比,因此管徑增加到一定程度後,若長度沒有隨之增加,則壁孔將成為限制輸水效率的因子,使輸水效率無法進一步上升。而管胞的管徑與長度也受細胞體積大小的限制而不能繼續增加[29],相較之下導管為多細胞的結構,其管徑與長度皆大於管胞,輸水效率也大幅提升[18]。被子植物的導管細胞很可能是由梯紋的管胞演化而來[5],即管胞上下兩側壁孔的壁孔膜漸消失,形成具有許多條狀構造的原始穿孔板(perforation plate)[30]。另外少數缺乏導管的被子植物只以管胞輸送水分,且數個管胞間壁孔的通透性較高,因而形成功能類似導管的結構以提升輸水效率[8][31]。
壁孔的演化 编辑
管胞表面讓水分在細胞間流動的壁孔可能是聚生的原生質絲演化而來,以提升水分在管胞間與其他組織間流動的效率,但隨著壁孔大小的增加,對管胞結構穩定性的影響越大,並增加氣穴栓塞從相鄰細胞中侵入的風險,故壁孔的演化過程為平衡這兩個因素的結果。S型管胞的孔洞較小,但因木質化的疏水部分也具有孔洞,會降低水分子與管壁的附著力,使氣穴進入所需的壓力(air-seeding pressure)降低而更易進入;G型管胞的孔洞較大,不過其木質化的疏水部分不具有孔洞,以降低氣穴進入的機會;P型管胞的壁孔已與現生植物的類似,可能具有可能類似現生植物的佈滿小孔且未有木質化的壁孔膜(pit membrane),因其為親水材料,使氣穴難以進入,且當壁孔一側出現氣泡而產生壓力差時,壁孔膜可被擠壓往另一側的壁孔緣(pit border)。後來多種壁孔結構的演化使其在不降低管胞結構穩定性與增加氣穴進入機會的同時,進一步提升水分在壁孔中流動的效率,例如許多裸子植物與部分被子植物中的多次演化而來的具緣壁孔,與買麻藤和多支被子植物中多次演化而來的附物壁孔(vestured pit)[18]。
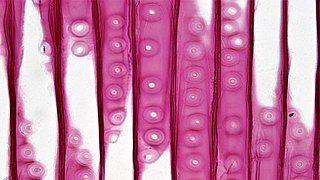
圖集 编辑
-
 歐洲蕨維管束的橫切面,經染色後,中間紅色的大型細胞即為管胞
歐洲蕨維管束的橫切面,經染色後,中間紅色的大型細胞即為管胞 -

-

-
 松樹樹幹的縱切,管胞上可見具緣壁孔
松樹樹幹的縱切,管胞上可見具緣壁孔

.jpg)
.jpg)
.jpg)
注释 编辑
- ^ 有些蕨類植物的管胞直徑可超過100微米[3]。
- ^ 包括星鱗蕨屬、苹屬、蕨屬與岩蕨屬等類群,因上下細胞間的細胞壁已幾乎消失,僅存一些細長的條狀殘跡,已符合導管定義[3]。
- ^ 買麻藤門為一例外,其為唯一一類具有導管的裸子植物[5],過去認為買麻藤門是與被子植物親緣關係最近的裸子植物,兩者合稱生花植物,但有許多研究不支持此假說,認為兩者為平行演化而來,故兩者木質部中的導管也是分別由不同類型的管胞演化而來[6][7]。
- ^ 根據哈根-帕醉方程,導管與管胞輸水的效率與其直徑的四次方成正比[9]。
- ^ 部分買麻藤與部分被子植物的管胞具有附物壁孔(vestured pit)[17],其壁孔腔中有支撐壁孔膜的木質化結構,使壁孔膜無法延展而避免氣穴的進入,但會降低水分在壁孔中流動的效率[18]。
参考文献 编辑
- ^ 假導管;管胞 tracheid. 國家教育研究院. 2012 [2019-06-04]. (原始内容存档于2019-06-04).
- ^ Sanio, C. Vergleichende Untersuchungen über die Elementarorgane des Holzkörpers. Bot. Zeitung. 1863, 21: 85–91; 93–98; 101–111. ISSN 2509-5420.
- ^ 3.0 3.1 Klaus Mehltreter, Lawrence R. Walker, Joanne M. Sharpe. Fern Ecology. Cambridge University Press. 2010: 147-148 [2019-05-29]. ISBN 113948768X. (原始内容存档于2022-01-06).
- ^ 4.0 4.1 R. Bruce Hoadley. Understanding Wood: A Craftsman's Guide to Wood Technology. Taunton Press. 2000: 18 [2019-05-29]. ISBN 1561583588. (原始内容存档于2021-11-17).
- ^ 5.0 5.1 Gurcharan Singh. Plant Systematics: An Integrated Approach. Science Publishers. 2004: 246 [2019-05-29]. ISBN 1578083516. (原始内容存档于2022-01-06).
- ^ Bowe, L. M.; Coat, G.; dePamphilis, C. W. Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales' closest relatives are conifers. Proceedings of the National Academy of Sciences. 2000, 97 (8): 4092–4097. ISSN 0027-8424. doi:10.1073/pnas.97.8.4092.
- ^ Chaw, S.-M.; Parkinson, C. L.; Cheng, Y.; Vincent, T. M.; Palmer, J. D. Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers. Proceedings of the National Academy of Sciences. 2000, 97 (8): 4086–4091. ISSN 0027-8424. doi:10.1073/pnas.97.8.4086.
- ^ 8.0 8.1 8.2 8.3 8.4 Choat, Brendan; Cobb, Alexander R.; Jansen, Steven. Structure and function of bordered pits: new discoveries and impacts on whole-plant hydraulic function. New Phytologist. 2008, 177 (3): 608–626. ISSN 0028-646X. doi:10.1111/j.1469-8137.2007.02317.x.
- ^ 9.0 9.1 Hans Lambers, F Stuart Chapin III, Thijs L. Pons. Plant Physiological Ecology. Springer Science & Business Media. 2008: 186 [2019-06-04]. ISBN 0387783415. (原始内容存档于2022-01-06).
- ^ Nardini, A.; Salleo, S.; Jansen, S. More than just a vulnerable pipeline: xylem physiology in the light of ion-mediated regulation of plant water transport. Journal of Experimental Botany. 2011, 62 (14): 4701–4718. ISSN 0022-0957. doi:10.1093/jxb/err208.
- ^ Esau, K. Anatomy of Seed Plants 2nd. New York: John Wiley and Sons. 1977. ISBN 0-471-24520-8.
- ^ Cecie Starr, Christine Evers, Lisa Starr. Biology: Concepts and Applications 10. Cengage Learning. 2016: 446 [2019-05-29]. ISBN 130596733X. (原始内容存档于2022-01-06).
- ^ Peter A. Raven; Ray F. Evert; Susan E. Eichhorn. Biology of Plants. W.H. Freeman and Company. 1999: 576–577. ISBN 1-57259-611-2.
- ^ Bollhoner, B.; Prestele, J.; Tuominen, H. Xylem cell death: emerging understanding of regulation and function. {Journal of Experimental Botany. 2012, 63 (3): 1081–1094. ISSN 0022-0957. doi:10.1093/jxb/err438.
- ^ James D. Mauseth. Botany. Jones & Bartlett Learning. 2016: 127 [2019-05-29]. ISBN 1284123588. (原始内容存档于2022-01-06).
- ^ Taylor, T. Introduction to Vascular Plant Morphology and Anatomy. Paleobotany (Second Edition) The Biology and Evolution of Fossil Plants. Academic Press. 2009: 201–222. doi:10.1016/B978-0-12-373972-8.00007-3.
- ^ Steven Jansen, Pieter Baas, Peter Gasson, Frederic Lens, and Erik Smets. Variation in xylem structure from tropics to tundra: Evidence from vestured pits. Proceedings of the National Academy of Sciences: 8833-8837. doi:10.1073/pnas.0402621101.
- ^ 18.0 18.1 18.2 18.3 18.4 18.5 18.6 Sperry, John S. Evolution of Water Transport and Xylem Structure. International Journal of Plant Sciences. 2003, 164 (S3): S115–S127. ISSN 1058-5893. doi:10.1086/368398.
- ^ 滕詠延. 管胞. 中華百科全書. 1983.[失效連結]
- ^ 20.0 20.1 Takayoshi Higuchi. Biosynthesis and biodegradation of wood components. Elsevier. 2012-12-02: 9 [2019-06-25]. ISBN 978-0-323-14328-8. (原始内容存档于2021-04-27).
- ^ John C.F. Walker. Primary Wood Processing: Principles and Practice. Springer Science & Business Media. 2006-09-13: 8 [2019-06-25]. ISBN 978-1-4020-4393-2. (原始内容存档于2021-04-27).
- ^ Paul G. Mahlberg. Laboratory Program in Plant Anatomy. Wm. C. Brown. 1972: 192 [2019-06-25]. (原始内容存档于2021-04-27).
- ^ T.T. Kozlowski. Seed Germination, Ontogeny, and Shoot Growth. Elsevier Science. 2012-12-02: 19 [2019-06-25]. ISBN 978-0-323-14947-1. (原始内容存档于2021-04-27).
- ^ 24.0 24.1 Marja-Sisko Ilvessalo-Pfäffli. Fiber Atlas: Identification of Papermaking Fibers. Springer Science & Business Media. 2013-03-09 [2019-06-25]. ISBN 978-3-662-07212-7. (原始内容存档于2021-04-27).
- ^ Samuel J. Record. Significance of Resinous Tracheids. John Merle Coulter, M.S. Coulter, Charles Reid Barnes, Joseph Charles Arthur (编). Botanical Gazette. University of Chicago Press. 1918: 61 [2019-06-25]. (原始内容存档于2021-04-27).
- ^ Sherwin Carlquist. Vasicentric Tracheids as a Drought Survival Mechanism in the Woody Flora of Southern California and Similar Regions; Review of Vasicentric Tracheids. A Journal of Systematic and Evolutionary Botany. 1985, 11 (1): 37-68. doi:10.5642/aliso.19851101.05.
- ^ Friedman, William E.; Edwards, D.; Read, D. J.; Duckett, J. G.; Cook, Martha E. The origin and early evolution of tracheids in vascular plants: integration of palaeobotanical and neobotanical data. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences. 2000, 355 (1398): 857–868. ISSN 1471-2970. doi:10.1098/rstb.2000.0620.
- ^ Paul Kenrick and Peter R. Crane. The Origin and Early Diversification of Land Plants: A Cladistic Study. Smithsonian Institution Press. 1997. ISBN 1-56098-730-8.
- ^ Lancashire, J.R. Modelling the hydrodynamic resistance of bordered pits. Journal of Experimental Botany. 2002, 53 (373): 1485–1493. ISSN 1460-2431. doi:10.1093/jexbot/53.373.1485.
- ^ Lens, Frederic; Vos, Rutger A.; Charrier, Guillaume; van der Niet, Timo; Merckx, Vincent; Baas, Pieter; Aguirre Gutierrez, Jesus; Jacobs, Bart; Chacon Dória, Larissa; Smets, Erik; Delzon, Sylvain; Janssens, Steven B. Scalariform-to-simple transition in vessel perforation plates triggered by differences in climate during the evolution of Adoxaceae. Annals of Botany. 2016, 118 (5): 1043–1056. ISSN 0305-7364. doi:10.1093/aob/mcw151.
- ^ Hacke, U. G.; Sperry, J. S.; Feild, T. S.; Sano, Y.; Sikkema, E. H.; Pittermann, J. Water Transport in Vesselless Angiosperms: Conducting Efficiency and Cavitation Safety. International Journal of Plant Sciences. 2007, 168 (8): 1113–1126. ISSN 1058-5893. doi:10.1086/520724.